Пиленгас - Mugil soiuy
Basilewsky, 1855
Популяционные характеристики
Популяции
пиленгаса из водоёмов нативного ареала и
водоёмов, в которых он акклиматизирован (Азово-Черноморский
бассейн) достаточно существенно отличаются
многими популяционными показателями (темпами
роста, размерами, временем полового
созревания и т.п.).
Для
дальневосточного пиленгаса характерны
невысокий темп роста, небольшие размеры и
более позднее половое созревание, чем у
пиленгаса из Азовского и Чёрного морей. Так,
пиленгас из Амурского залива созревает в
возрасте трех (самцы) -
четырех (самки) лет при достижении длины 28-30
см. В возрасте 10 лет дальневосточный
пиленгас достигает массы 2700 г. Более
крупные экземпляры этого вида в морях
дальнего Востока не известны (Мизоркина А.В.
и др., 1981).
В Азово-Черноморском регионе
темпы роста и размеры пиленгаса
значительно больше. По данным Ю.В. Пряхина
(1996), средние показатели массы тела
двухгодовиков пиленгаса из Азовского моря
составляют 640 г, трехгодовиков -
954 г, четырехгодовиков -
1341 г, пятигодовиков -
2572 г, шестигодовиков -
2996 г. В азовских лиманах пяти- и шестилетки
имеют длину 60-65
см и массу 3,5-5,0
кг.
Далее в
сравнительном аспекте опишем размеры,
упитанность и индексы печени пиленгаса из
Молочного лимана Азовского моря и из
Кизилташских лиманов, связанных с Чёрным
морем.
В Молочном
лимане Азовского моря в уловах нередки
особи длиной более 80 см и массой 6 кг. В
системе Кизилташских лиманов, активное
заселение которых пиленгасом началось с
конца 80-х гг. ХХ века, а естественный нерест
впервые отмечен в начале 90-х гг., стадо
пиленгаса еще достаточно разрежено.
Половозрелые особи в этих водоемах имеют
среднюю длину 45-55
см при массе 1000-1500
г. По-видимому, когда массовое потомство
особей пиленгаса, отнерестившихся в
Кизилташских лиманах, достигнет возраста 7-8
лет, крупные особи будут встречаться в
уловах значительно чаще (Глубоков, Микодина,
1996).
А.И. Глубоков и Е.В.
Микодина (1996) сообщают, что коэффициенты
упитанности половозрелых особей пиленгаса
сходны у азовского и черноморского стад.
Эти показатели в течение года колеблются в
пределах 1,06-1,39.
Наивысших значений коэффициенты
упитанности достигают перед нерестом и в
конце нагульного периода, а минимальных –
сразу после нереста и после зимовки.
Коэффициенты упитанности самцов ниже, чем
самок в обоих изученных стадах, хотя самцы,
в отличие от самок, запасают больше
количество жира в полости тела и на
кишечнике. Единственным отличием по
данному показателю азовского пиленгаса от
черноморского является большее снижение
коэффициентов упитанности после нереста у
особей азовского стада. Так, в начале июля
средние коэффициенты упитанности особей
пиленгаса из Молочного лимана составляют
1,06-1,07,
а из Кизилташских лиманов – 1,14-1,15.
Это свидетельствует о более полной
реализации репродуктивных возможностей
стадом из Молочного лимана по сравнению с
Кизилташским стадом. Отмеченная
особенность, по-видимому, связана со
степенью соответствия в этих водоемах
условий нереста и нагула оптимальным
параметрам (Глубоков, Микодина, 1996).
Динамика
колебаний индексов печени у особей из
Молочного и Кизилташских лиманов сходна.
Поскольку печень является депо запасных
питательных веществ, наибольших значений
(2,9-3,1)
индексы печени достигают в конце нагула – в
октябре. Перед нерестом этот показатель
также имеет высокие значения – 2,6-3,2,
а после его завершения расход энергии столь
велик, что относительный размер печени
становится минимальным: индексы печени в
начале июля колеблются в пределах 1,2-1,4.
Исследование в Кизилташских лиманах
годовой динамики индексов печени у рыб, не
достигших половой зрелости, показало
сходство характера изменений этого
показателя с половозрелыми особями (Глубоков,
Микодина, 1996).
Анализируя
полученные данные, А.И. Глубоков и Е.В.
Микодина (1996) пришли к выводу, что основные
морфофизиологические показатели пиленгаса
Кизилташских лиманов существенно не
отличаются от таковых пиленгаса из
азовского стада. Специфические условия
Черноморского бассейна не привели к
формированию устойчивых адаптивных
изменений, выявляемых с помощью
морфофизиологических показателей, что
позволяет в настоящее время относить оба
стада к одной популяции.
Экология
Пиленгас
относится к рыбам эстуарно-морского типа.
Являясь эвригалинной рыбой, он может
обитать на всех этапах постэмбрионального
развития как в пресной воде, так и в водах с
морской и океанической соленостью (Казанский,
1989).
В зависимости
от времени года пиленгас встречается в
различных биотопах. Так, Б.Н. Казанский (1989)
сообщает, что на Дальнем Востоке пиленгас
на зимовку заходит в реки и их низовья, где
залегает в ямы на глубине 6-10
м. По р. Раздольная он поднимается на 100 км
вверх по течению и держится там 100-120
дней. Заход пиленгаса в реки на зимовку
происходит в конце октября -
начале ноября при резком похолодании, ход
довольно дружный. После зимовки, в конце
марта -
начале апреля, пиленгас выходит из рек в
воды Амурского залива, распределяется на
илистых мелководьях.
Иначе обстоит
дело с распределением азовского пиленгаса,
который высокоактивен практически в
течение всего года. Ю.В. Пряхин с соавторами
(2000) приводят данные, согласно которым
сеголетки и часть наиболее мелких
двухлеток обычно заходят на зимовку в
пресные водотоки. В отличие от устоявшегося
мнения, основанного на данных по водоемам
Дальнего Востока, молодь пиленгаса даже при
температуре воды около 2°С
не держится постоянно в устойчивых
скоплениях, а временно рассредоточивается
или концентрируется, способна перемещаться
с одного участка зимовки на другой и
развивать высокие бросковые скорости в
случае опасности.
Молодь не
только мигрирует по пресным водотокам, но и
кратковременно выходит в прибрежные районы
моря и способна перемещаться из одного
водотока в другой. При наличии прибрежного
льда сеголетки днём распределены
относительно разреженно под ним или
отстаиваются на ямах. В сумеречное время
происходит выход рыбы на центральные
участки русла. При оттепелях в ясные дни
скопления сеголеток часто не распадаются, а
если перемещаются, то обычно по направлению
ветра. В скоплениях наблюдается эффект
броуновского движения: в течение
относительно короткого промежутка времени
происходит изменение формы косяков,
разделение их на более мелкие или
воссоединение (Пряхин и др., 2000).
Исследователи
пишут, что большая часть двух и трехлеток
зимует в прибрежье Азовского моря и в
отдельных лиманах. Основная часть
старшевозрастных особей распределяется в
открытых районах центральной и западной
частей Азовского моря. В восточной, имеющей
более обширную мелководную зону,
значительных зимовальных скоплений
крупного пиленгаса не обнаружено (Пряхин и
др., 2000).
Установлено, что пиленгас
реагирует не только не резкие колебания
температуры воды, но и на тенденции к ее
изменению. Даже при слабом росте
температуры воды в зимний период
активность рыбы повышается, а скопления
становятся менее устойчивыми. Кроме этого,
распределение пиленгаса в течение зимовки
непостоянно и может меняться под
воздействием складывающихся температурных
и гидрологических условий (Пряхин и др., 2000).
Важной
особенностью биологии пиленгаса Азово-Черноморского
бассейна является ежегодная массовая
миграция в весенний период части его
производителей через Керченский пролив в
Черное море. Достоверной оценки количества
особей, покидающих Азовское море, еще пока
нет, но масштабы этого явления значительны.
Есть мнение, что в Черное море мигрирует
большая часть нерестовой популяции. Таким
образом, происходит постоянное пополнение
черноморского стада. Закономерности,
влияющие на интенсивность этого процесса и
количество мигрирующих в Черное море рыб
пока не установлены (Пряхин и др., 2000).
Размножение
Нерест
пиленгаса на Дальнем Востоке происходит со
второй половины мая до первой половины июля
при температуре воды 18-24°С.
Он растянут за счет разного времени
подходов для нереста и, возможно,
двухпорционности икрометания. Самцы
созревают в возрасте 4, а самки – 5 лет.
Нерест происходит у берегов над небольшими
глубинами, икра пелагическая. Нерест
происходит довольно дружно, в нем с одной
крупной самкой участвуют несколько самцов.
Пиленгас при нересте избегает океанических
соленостей и избирает воду с солёностью 18-26%0,
возможно, может нереститься и при
большей опресненности. В опытах особенно
длительная подвижность спермиев
отмечалась в морской воде примерно при 16%0.
Однако и в воде при 32-33%0
активность всех спермиев была высокой,
хотя время их подвижности сокращалось до 11-18
минут (Казанский, 1989).
На зимовку в
пресные водоёмы Дальнего Востока пиленгас
заходит в конце октября -
ноябре с незавершенным гаметогенезом.
Видимо, в период зимовки, когда пиленгас
прекращает питаться, гаметогенез у него
сильно тормозится. Вода во многих
быстротекущих реках предгорий Сихоте-Алиня
зимой значительно охлаждается – до 0°С.
В это время в течение 100-120
суток основная трата на энергетический
обмен идет за счет мускулатуры, жирность
которой снижается с 10-13%
до 4%, но при этом почти не расходуется
внутриполостной жир (Казанский, 1989).
В марте -
апреле пиленгас выходит из рек и усиленно
питается на мелководьях. В это время
гаметогенез у него сильно активизируется. К
началу нерестовой миграции в середине мая
гонады самцов находятся еще в состоянии
активного сперматогенеза, а в семенных
ампулах присутствуют клетки всех фаз
развития. К концу мая количество более
зрелых самцов возрастает, однако этот
процесс идет очень асинхронно. Видимо,
поэтому самцы подходят на нерест не все
сразу и участие в нем растянуто во времени (Казанский,
1989).
У созревающих
самок при выходе с зимовальных ям гонады
находятся в III -
начале IV
стадии зрелости, ГСИ варьирует от 8 до 15%. В
завершенной IV
стадии зрелости, когда яичники достигают
массы 200-380
г, ГСИ достигает максимального значения -
22%. Примерно за один весенний месяц ГСИ
увеличивается в 1,5-3
раза. Этот процесс у самок также происходит
асинхронно, и их участие в нересте
растягивается (Казанский Б.Н., 1989).
Не вполне ясен
вопрос о типе икрометания пиленгаса ввиду
наличия у самок IV стадии зрелости двух групп
желтковых ооцитов – в фазе отложения
глубоководного желтка и в фазе
наполненного желтком ооцита. Наряду с
растянутостью нерестового сезона, можно
предположить двухпорционность икрометания.
Однако судьба менее развитых желтковых
ооцитов может быть различной. Как показали
наблюдения при выращивании пиленгаса в
бассейнах, такие ооциты резорбируются.
Вполне вероятно, что у пиленгаса имеется
возможность двухпорционного икрометания,
которая реализуется лишь в наиболее
благоприятных условиях (Казанский, 1989).
Сведения по
размножению пиленгаса в Азово-Черноморском
регионе следующие.
По данным Е.Б.
Моисеевой и А.К. Любомудрова (1997), семенники
азово-черноморского пиленгаса
представляют собой парные органы
мешковидной формы, расположенные по обеим
сторонам кишечника и плавательного пузыря.
Снаружи они покрыты тонкой эластичной
оболочкой. У молодых рыб поверхность
семенников ровная, у более взрослых
образует несколько складок, разделяющих
гонады на крупные доли. Часто один семенник
бывает заметно крупнее другого, иногда
встречаются особи с одной железой. В
зависимости от функционального состояния и
степени зрелости семенников цвет, форма и
размеры их существенно меняются.
Помимо мужских
половых клеток в семенниках многих самцов
пиленгаса встречаются половые клетки
противоположного пола. Е.Б. Моисеевой и А.К.
Любомудровым (1997) они были обнаружены у 45,7%
исследованных рыб. Овоидные клетки
встречаются в семенниках во все сезоны года,
а их число варьирует от единичных клеток в 20-80
срезах до сотни в одном срезе. Самцы с
большим числом ооцитов встречаются в
основном в июле и сентябре. Авторы считают,
что развитие ооцитов в семенниках не идёт
далее начальных стадий периода
протоплазматического роста (Моисеева Е. Б.,
Любомудров А. К., 1997).
На основании
анализа семенников пиленгаса можно
представить схему годичного полового цикла
самцов этого вида в Азово-Черноморском
бассейне. Наиболее длительной является
вторая стадия зрелости, продолжающаяся с
августа по февраль-март. Период размножения
сперматогониев протекает в широком
диапазоне температуры, от 24°С
в августе до 2,9°С
в ноябре. В зимние месяцы размножение
сперматогониев, по-видимому,
приостанавливается, возобновляясь при
поднятии температуры до 2,2-6,0°С
в феврале-марте.
Стадии зрелости
II-III
и III,
отражающие начало и конец периода
созревания половых клеток, длятся с января
по апрель -
начало мая. Температура воды, на фоне
которой проходят эти стадии, изменяется от 5°С
в январе до 16°С
в мае. Процесс созревания половых клеток
наиболее дружно протекает на фоне
температуры, повышающейся с 9,6-11°С
(апрель) до 15,5-16,8°С
(май). Формирование сперматозоидов,
наступающее вслед за началом созревания и
идущее параллельно с ним, завершается за
один -
два месяца и в том же температурном
диапазоне, что и конечные этапы созревания
(10-17°С).
К концу мая у большинства самцов гонады
находятся на IV
стадии зрелости. Нерест рыб длится с
середины мая по середину июня, при
температуре воды 20,5-23,5°С.
К середине -
концу июня большинство самцов завершают
годичный цикл размножения. Процессы
резорбции остаточных сперматозоидов и
восстановление обычной структуры желез
длятся один -
полтора месяца, с конца июня по начало
августа, на фоне температуры, варьирующей в
пределах 23-26
°С
(Моисеева, Любомудров, 1997).
В пределах
Азово-Черноморского бассейна участки
поверхностного слоя водной толщи с
соленостью свыше 18%0 весьма ограничены. Поэтому
возможность естественного воспроизводства
пиленгаса в солоноватоводном Азовском море
длительное время оспаривалась и
представлялась реальной лишь в небольших
объемах на ограниченных площадях отдельных
придаточных водоемов. Пиленгаса
рассматривали прежде всего как
перспективный объект пастбищного и
товарного выращивания на базе заводского
воспроизводства (Воловик, 1999).
В начальный
период акклиматизации пиленгаса инкубацию
его икры проводили при солености 15-23%0
и температуре 17-22°С.
Было установлено, что обязательным
условием нормального развития икры
является взвешенное состояние, а
нейтральная плавучесть икры зависит от ее
размеров, массы, обводненности и солености
воды. Наиболее благоприятна для развития
крупной икры вода с соленостью 23-30%0
. Мелкая икра нормально развивалась в
менее соленой воде. Опыты показали, что икра
пиленгаса оплодотворяется в довольно
широком диапазоне солености – от 3-5
до 45%0 . В
пресной воде оплодотворения не происходит.
При этом в “спокойной” воде с соленостью
менее 20 и более 30%0 смертность возрастает, а при
слабой аэрации, позволяющей икре
находиться во взвешенном состоянии, выклев
и развитие личинок наблюдаются даже в
распресненной до 10-15%0
воде. В экспериментах с применением
аэрации выклев личинок варьировал от 65% (при
солёности 5%0)
до 82% (при солёности 25%0)
(Воловик, Пряхин, 1999).
В условиях
Азово-Черноморского бассейна у пиленгаса
было отмечено уменьшение размеров икры при
некотором увеличении объема жировой капли
и общей плодовитости особей.
Значительная
вариабельность морфо-экологических и
биохимических показателей икры пиленгаса (относительный
объем жировой капли, количество липидов,
плавучесть) играет важную роль в адаптации,
гарантирующей воспроизводство и выживание
пиленгаса в новых эколого-гидрологических
условиях Азово-Черноморского бассейна.
Уменьшение диаметра икры при одновременном
увеличении жировой капли повышает
плавучесть икры и обеспечивает нормальное
развитие во взвешенном состоянии при
относительно низкой солености. Эти
изменения – адаптация, направленная на
обеспечение более эффективного
использования нерестилищ за счет развития
икры не только в слое гипонейстона, но и в
пелагиали (Старушенко, 1998).
Приведенные
данные свидетельствуют о начале
формировании в Азово-Черноморском бассейне
неоднородной популяции пиленгаса,
способной использовать для
интенсивного и эффективного нереста
районы с большими отличиями по солености
воды. Учитывая опыты по аэрации в период
инкубации, можно допустить, что течения и
волнение моря способствуют поддержанию
икры во взвешенном состоянии даже при
относительно низкой солености. В природных
условиях перемешивание водной толщи в
придонном горизонте до глубины 15 м
происходит уже при скорости ветра 7 м/c.
В период размножения пиленгаса и развития
его икры такие ситуации возникают более чем
в 90% случаев (Пряхин, 1999). Относительно
сроков нереста азово-черноморского
пиленгаса мнения исследователей
разделились. Так, Т.П. Чесалина (1997) считает,
что в Черном море пиленгас размножается с
конца мая до середины июля. С.П. Воловик и Ю.В.
Пряхин (1999) сообщают, что во второй половине
июня 1998 г. в открытой части Азовского моря 60%
самок и 40% самцов пиленгаса имели гонады в
стадии выбоя, 28% самок и 26% самцов были с
созревающими или зрелыми гонадами, у 11%
самок размерами 30-50
см и 34% самцов размерами 31-55
см они были в состоянии покоя.
А.И. Глубоков и Е.В.
Микодина (1996) приводят данные, согласно
которым в Кизилташских лиманах нерест
пиленгаса наступает в конце мая -
начале июня, а в Молочном лимане Азовского
моря -
несколько раньше, в начале мая. Более
поздние сроки нереста черноморского стада
пиленгаса по сравнению с азовским
обусловлены, по мнению авторов, более
медленным прогревом воды в Черном море.
Гонадо-соматические
индексы (ГСИ), по которым можно судить о
степени готовности рыб к нересту, у самцов
пиленгаса из Молочного лимана достигают
максимальных значений во второй половине
мая и составляют в среднем 10,7%. У самок
азовского пиленгаса этот показатель перед
нерестом составляет в среднем 16,8%.
Минимальные значения ГСИ у азовского стада
отмечены после полного завершения нереста
– в начале августа. Их величина у
половозрелых особей составляет в среднем
0,3% у самцов и 0,9% у самок. У особей
кизилташского стада, участвующих в нересте,
максимальные величины ГСИ несколько ниже, а
минимальные соответствуют таковым у особей
из Молочного лимана. Они составляют 9,0-12,6%
и 0,3% соответственно. Таким образом, ГСИ
демонстрирует большую степень реализации
репродуктивного потенциала азовским пиленгасом
по сравнению с черноморским (Глубоков А.И.,
Микодина Е.В., 1996).
Относительно
динамики ГСИ в процессе полового
созревания особей из системы Кизилташских
лиманов можно отметить следующее. На втором
году жизни ГСИ составляет 0,02%, на третьем он
возрастает с 0,07% в начале вегетационного
периода до 0,33% в его конце. На четвертом году
большая часть самцов и некоторые самки
впервые вступают в нерест (Глубоков,
Микодина, 1996).С.В.
Пьянова (2002) также обнаружила, что значения
ГСИ выше у производителей пиленгаса из
Азовского моря. Первые
случаи обнаружения оплодотворенной икры и
личинок пиленгаса азовской популяции были
отмечены в придаточных морских водоемах с
повышенной минерализацией воды (13-18%0)
– в Молочном и Сивашском лиманах. Позднее
икра и личинки были отмечены в Ханском
озере, Бейсугском и ряде других лиманов
Сладковской, Ордынской и Кизилташской
систем кавказского побережья (Воловик,
Пряхин, 1999).
Ежегодные
ихтиопланктонные съемки показали, что
пиленгас освоил в качестве нерестового
ареала большую часть акватории Азовского
моря с соленостью свыше 8%0 . Появление
высокоурожайного поколения 1994 г.
сопровождалось распределением икры
пиленгаса на акватории 13,2 тыс. км2 .
Максимальные уловы достигали 22 икринок и 80
личинок на один облов икорной сети ИКС – 80.
Расчетная численность икры составила около
194 млн. штук, а личинок – 672 млн. штук (Воловик,
Пряхин, 1999).
В июне 1996 г.
впервые была обнаружена нормально
развивающаяся икра пиленгаса в Черном море
в районе Севастополя (Чесалина, 1997).
Икринки
пиленгаса сферической формы, оболочка их
гладкая, тонкая и прозрачная, желток
гомогенный светло-желтого цвета. Внутри
икринки располагается одна крупная жировая
капля (0,36-0,40
мм) (рисунок). Икринки пелагические, плавают
у самой поверхностной пленки воды жировой
каплей кверху. Осемененные икринки
начинают набухать, через 10-20
минут от момента оплодотворения между
желтком и оболочкой появляется
перивителлиновое пространство,
составляющее 3,3-16,6%
диаметра икринки.
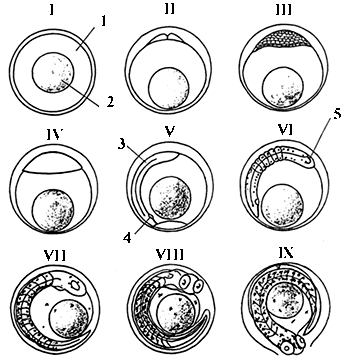
Икра пиленгаса на разных стадиях
развития (по Чесалиной, Чесалину, 1997).
Через 40 минут -
1 час после оплодотворения на анимальном
полюсе икринки формируется
цитоплазматический бугорок, который
начинает делиться и образовывать
бластомеры.
Примерно через 1
час 30 минут появляется первая борозда
дробления, а через 2 часа 10 минут -2
часа 20 минут уже образуются 4 бластомера;
через 4 часа 30 минут -
4 часа 40 минут – 32. Спустя 4 часа 50 минут
образуется морула крупных клеток, через 6
часов – средних, через 6 часов 50 минут -
7 часов – мелкоклеточная морула. Процесс
дробления бластомеров приводит к
формированию куполообразного дисковидного
скопления клеток на анимальном полюсе
икринки – бластодермы. Весь процесс
дробления при температуре воды 20,5-21°С
продолжается около 6 часов (Чесалина,
Чесалин, 2000).
В возрасте
около 7 часов после оплодотворения купол
бластодермы начинает постепенно
уплощаться. При повороте икринки внутри
бластодермы хорошо заметна полость
бластоцеля. Продолжительность этапа
бластулы составляет 4 часа.
Через 11 часов -
11 часов 30 минут с момента оплодотворения
начинается процесс эпиболии – бластодерма
нарастает на желток. В возрасте эмбриона 21-22
часа процесс обрастания желтка
заканчивается. К этому времени в икре
хорошо просматривается зародышевая
полоска шириной 0,10-0,12
мм. Головной конец ее расширен, начинается
закладка хорды. В центральной части
зародышевой полоски появляются мелкие
точечные меланофоры. Гаструляция при
температуре 21-21,5°С
продолжается 10-11
часов (Чесалина, Чесалин, 2000).
В возрасте 22
часа тело зародыша располагается на
желточном мешке в виде валика. Хорошо
заметна хорда вдоль его оси. В головном
отделе есть зачатки глазных бокалов и
мозговых пузырей. Отмечена сегментация
туловищного отдела. В возрасте 25 часов у
эмбрионов четко обозначены глазные бокалы,
сформированы мозговые пузыри, число
туловищных сомитов достигает 10-11.
Тело пигментировано точечными
меланофорами, располагающимися двумя
рядами вдоль спинной стороны туловищного
отдела. Длительность этого этапа
составляет около 6 часов.
В возрасте
около 28 часов начинается отделение
хвостового отдела зародыша от желточного
мешка и его рост. Вместе с ростом хвостового
отдела идет увеличение длины хорды,
нервного тяжа, формируются новые хвостовые
сегменты. Через 30 часов с момента
оплодотворения эмбрион охватывает 2/3
желтка, в теле насчитывается до 20-22
сегментов, есть зачатки сердца, печени,
кишечника, плавниковой каймы. Число мелких
пигментных клеток на теле увеличивается,
окраска их становится более интенсивной, на
желтке и жировой капле появляются
звездчатые меланофоры. В возрасте 32 часов
звездчатые меланофоры образуются на голове
и спине. Кроме этого, на голове и брюшной
стороне появляются пигментные клетки желто-коричневого
цвета.
В возрасте 35-36
часов эмбрион становится подвижным, у него
начинает пульсировать сердце. Кровь почти
бесцветна. В теле насчитывается 24 миомера.
Голова плотно прижата к желточному мешку, а
хвост достигает головы. Тело зародыша
окружает плавниковая кайма. Эмбрион (кроме
задней половины хвостового отдела)
интенсивно пигментирован. Зародыш свободно
вращается внутри оболочки, резкими
движениями прорывая оболочку и
освобождаясь от нее. Длительность этапа при
температуре 22°С
-
6-7
часов.
В целом
эмбриональное развитие пиленгаса внутри
оболочки при температуре воды 20,5-22°С
продолжается 41-43
часа. Продолжительность процесса
эмбриогенеза зависит от температуры
инкубации. Так, период инкубации икры
пиленгаса при понижением температуры воды
с 23°С
до 17°С
удлиняется с 36 до 60 часов (Чесалина, Чесалин,
2000).
Только что
выклюнувшиеся предличинки пиленгаса имеют
длину 2,4-2,6
мм. Тело их короткое и высокое. У них
сохраняется большой желточный мешок длиной
около 0,67 мм с крупной жировой каплей
диаметром 0,42-0,44
мм. Голова плотно прижата к желточному
мешку, ротовое отверстие отсутствует. В
туловищном отделе содержится 10-11,
в хвостовом – 13-14
миомеров. Плавниковая кайма хорошо
сформирована, начинается от головного
отдела и тянется до ануса. Антеанальное
расстояние в среднем составляет 58% от длины
тела (рисунок):
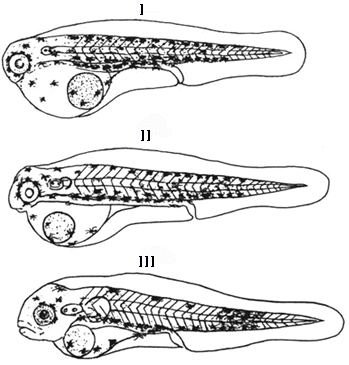
I
– только что вылупившаяся предличинка; II
– предличинка в возрасте 1 сутки; III -
предличинка в возрасте 3 суток
Предличинки пиленгаса
на разных стадиях развития (по Чесалиной,
Чесалину, 2000).
Сердце
двухкамерное, число его сокращений
составляет 140-150
ударов в минуту, кровь не окрашена. Имеются
зачатки грудных плавников в виде бугорков.
Глаза не пигментированы. На голове вокруг
глаз и на темени расположены единичные
меланофоры. Туловищный отдел и начало
хвостового сильно пигментированы
точечными и звездчатыми меланофорами.
Задняя часть хвостового отдела, примерно с
его середины лишена пигмента. На желтке и
жировой капле имеются единичные звездчатые
меланофоры. Верхняя часть кишечника сильно
пигментирована меланофорами. Помимо
черного пигмента есть пигментные капли
желто-коричневого цвета, которые
сосредоточены преимущественно на брюшной
стороне тела, а также на голове и спинной
стороне, окрашивая эти участки в желтый
цвет.
Через сутки
после выклева длина предличинок достигает
2,65-2,90
мм. Длина желточного мешка уменьшается до
0,59 мм. Есть небольшие грудные плавники.
Число миомеров в сумме составляет 24. Глаза
не пигментированы, пигментация тела
немного ослабляется. Более четко
выделяются черные пигментные клетки по
брюшной стороне, а на спине пигмент
светлеет. На голове хорошо просматривается
несколько звездчатых меланофоров.
На третьи сутки
предличинки достигают длины 2,91-3,30
мм. У них просматривается желточный мешок
длиной около 0,48 мм. Открывается конечный
рот. В глазах появляется черный пигмент.
Антеанальное расстояние уменьшается до 55%
длины тела. Грудные плавники увеличиваются.
В крови появляются единичные эритроциты. На
голове располагаются отдельные крупные
звездчатые меланофоры. Пигментация
туловищного отдела ослабевает, а за анусом
образуется широкая поперечная полоска из
большого числа меланофоров. Тело
предличинок окрашено в желтый цвет
клетками желто-коричневого цвета.
Звездчатыми меланофорами сильно
пигментирована верхняя часть кишечника.
Клетки желто-коричневого цвета
локализованы у основания грудных плавников
и на брюшной стороне предличинки.
Активность
предличинок невелика, лишь изредка они
совершают вертикальные перемещения в толще
воды (Чесалина, Чесалин, 2000).
Питание
Для пиленгаса,
как и для подавляющего большинства видов
рыб, характерна смена объектов питания в
онтогенезе. Первые два месяца после
перехода на внешнее питание личинки и
молодь питаются зоопланктоном. В этот
период рот у молоди конечный, затем он
становится нижним, а нижняя губа
приобретает форму скребка. В возрасте двух
месяцев пиленгас становится типичным
детритофагом и не является конкурентом
другим рыбам по зоопланктону и зообентосу.
Для
дальневосточного пиленгаса характерно
питание на илистых мелководьях. В желудках
преобладает детрит (до 80% по массе), в состав
которого входят преимущественно остатки
диатомовых и зеленых водорослей, в
незначительном количестве встречаются
амфиподы и другие животные, которые иногда
составляют до 6% по массе и, видимо,
заглатываются неизбирательною. Иногда до 10%
по массе составляет фитопланктон.
Пища, поступает
в ротовую полость, отфильтровывается на
жаберных тычинках, откуда направляется в
глотку, где уплотняется, после чего
перемещается в переднюю, сильно
растягивающуюся часть желудка. Отсюда пища
попадает в мускулистую часть желудка, где
частично перетирается, после чего
переходит в чрезвычайно длинный кишечник (Никольский,
1950).
Обычно индексы
наполнения желудка у пиленгаса выше, чем у
других видов рыб. Хотя в период нереста
встречается значительная часть самок с
пустыми желудками. Видимо в этот период
интенсивность их питания сильно снижается (Казанский,
1989).
В Азово-Черноморском
бассейне помимо детрита в желудках
пиленгаса встречаются полихеты, хирономиды,
молодь креветок, моллюск гидробия (Пряхин,
1996, 1997) и нитчатые водоросли. Индексы
наполнения желудков колеблются от 500 до 1240%00 (Демьяненко,
1999).
Осенью при
охлаждении воды до 5-8°С
пиленгас прекращает нагул и,
соответственно, питание (Старушенко Л. И.,
1989).
Значение
Пиленгас -
один из немногих видов рыб-акклиматизантов,
ставших объектами рыбного хозяйства (Пьянова,
2002). Причём его хозяйственное значение
более велико в водоёмах вселения, чем в
естественном ареале.
В водоемах
Дальнего Востока уловы пиленгаса невелики.
Так, по данным А.В. Мизюркиной и В.Г.
Марковцева (1981), добыча пиленгаса
в Южном Приморье лишь в отдельные годы
достигает 500 т. Причём в древности пиленгас
имел несомненно более существенное
хозяйственное значение. Л.Н. Беседнов
(1998) сообщает, что из 328 исследованных
костных остатков рыб неолитической
раковинной кучи в бухте Бойсмана залива
Петра Великого, датированной возрастом 6,5-5,0
тыс. лет, более 98% остатков принадлежит
пиленгасу. Средняя длина ископаемого
пиленгаса (47,3 см) превосходит таковую из
современных уловов на 15 см, что объясняется,
видимо, ухудшением условий обитания
дальневосточного пиленгаса в течение
последних тысячелетий (Беседнов Л.Н., 1998).
Гораздо большую
роль в уловах имеет азово-черноморская
популяция пиленгаса. По экспертным оценкам,
запас промысловой части азовской популяции
пиленгаса варьирует в пределах 30-45
тыс. т в начале и 20-30
тыс. т в конце зимовки. Официальный вылов
пиленгаса за зимний период 1998-1999
гг. составил порядка 7,5 тыс. т. Не менее
половины объёма официального составил
браконьерский вылов пиленгаса (Пряхин и др.,
2000).
Наиболее
благоприятны для промысла пиленгаса зимний
и весенний периоды, когда он образует
достаточно плотные зимовальные и
миграционные скопления, а активность рыбы
не так велика, как летом. В теплый период
года, вследствие природной осторожности,
подвижности, прыгучести рыбы и большой силы
броскового удара, лов пиленгаса
малоэффективен (Пряхин, 1996).
Основные виды
орудий лова, применяемые при промысле
пиленгаса, традиционны, так как практически
взяты из предшествующего опыта ловли
кефали: кефалевые ставные невода, ставные и
обкидные сети, различные модификации
закидных неводов и волокуш, а при обловах
зимних скоплений в море – кошельковые
невода и пелагические тралы (Пряхин, 1996).
Однако
особенности поведения пиленгаса снижают
эффективность применения того или иного
орудия лова. Так, В.В. Кузнецов (1998) указывает,
что у пиленгаса сформировалась устойчивая
оборонительная реакция на сетное полотно, а
Ю.В. Пряхин и др. (2000) считают трал
малоэффективным орудием лова пиленгаса в
Азовском море.
Специалисты
возлагают на пиленгаса большие надежды при
разработке проектов повышения
рыбопродуктивности Азово-Кубанских
лиманов. Потребляя почти не используемые
кормовые ресурсы детрита и давая высокие
среднегодовые приросты, пиленгас является
весьма перспективным объектом лиманного
рыбоводства (Цуникова и др., 2001).
Высокая
эврибионтность пиленгаса предопределила
возможность его выращивания в пресноводных
прудах в поликультуре с карпом и
растительноядными рыбами. Согласно данным
украинских специали
Источник: http://akklimatizant.narod.ru/mugil.htm |

